
لاله واژگون
لاله واژگون از منحصربفردترین گونه گلها و گیاهان بومی و وحشی مناطق کوهستانی ایران است. این گونه گیاهی در ارتفاعات سردسیر کهکیلویه و بویراحمد، اشترانکوه لرستان، کوههای صمصامی چهار محال و بختیاری و گلستان کوه در خوانسار، در سطح وسیع و قابل توجهی می روید، که به عقیده گیاه شناسان، این گل سربهزیر به سرعت در حال انقراض بوده و زیستگاه و تعداد این گونه گیاهی بسیار کم شده است.
این گونه مقاومت زیادی در برابر سرما و سازگاری ویژهای با دامنههای سنگلاخی و صخرهای دارد و چنانچه زیستگاه این گل به وسیله انسان و دام تخریب نشود، توانایی پوشاندن عرصههای وسیعی از طبیعت را با رویش هزاران شاخه زیبا دارد. گیاه لاله واژگون از جمله گیاهان در خطر انقراض کشور است که هرساله به دلیل برداشت غیر قابل کنترل گل و پیاز از رویشگاههای طبیعی، بیش از پیش در معرض نابودی قرار دارد. از دست دادن این نعمت بزرگ که از میلیونها سال پیش به ما به ارث رسیده است، ضایعهای بزرگ خواهد بود. با توجه به ضرورت حفظ این نوع گونههای بومی، در این پژوهش سعی شده است تا گامی هر چند کوچک در جهت روش ازدیاد به منظور حفظ لاله واژگون برداشته شود. امید است این کوشش بتواند گامی در راستای حفظ محیط زیست و منافع ملی تلقی شود. بدون تردید برداشتن گامهای بلند در آینده توسط متخصصان، در گرو بهرهگیری از یافته های کوچک خواهد بود. به امید آن روز.

در ایران لاله واژگون تحت تاثیر دمای هوا از اوایل بهار در نواحی شمالی زاگرس شروع به شکوفایی میکند و در مناطق جنوبی در اواسط تابستان به گل مینشیند. لاله واژگون تا ۱۰۰ سانتیمتر از سطح زمین ارتفاع دارد و عمر این گیاه بسیار کوتاه است .از اوایل اردیبهشت ماه گلدهی این گیاه شروع و در پایان فصل بارش تمام می شود. این گونه بوتهای در منطقه حفاظت شده مانشت (ارتفاعات شمال شهر ایلام) از اوایل اردیبهشت ماه تا اواخر خرداد ماه در دو رویشگاه تنگ دالاب و دامنه کوه مانشت رشد میکند.
در ایران براساس سیاستهای سازمان حفاظت محیط زیست، لاله واژگون به عنوان ذخیره ژنتیکی و عنصر زیبایی شناختی قلمداد میگردد و به همین دلیل حفاظت از آن ضروری است. با توجه به شرایط موجود (پراکنش محدود و متراکم) به اضافه چرای دامها، جاده سازی، بوتهکنی، برداشت گل و سوخ، قاچاق گل و عرضه به بازار، به نظر میرسد که بقای لاله واژگون در آینده با چالش روبهرو گردد. بر اساس مصوبه شماره ۱۵۰ مورخ ۲۷/۱۱/۷۵ شورای عالی محیط زیست، وضعیت لاله واژگون نگران کننده اعلام شده است.
شدت چرا دام ها در برخی مناطق به حدی است که گلدهی سال بعد را با مشکل مواجه میکند زیرا لاله واژگونکه اندام هوایی خود را سریع از دست میدهد از ذخیره غذایی کافی برخوردار نبوده و در نتیجه گل سال بعد از کیفیت چندانی برخوردار نخواهد بود و چنانچه در سالهای متوالی این تخریب صورت پذیرد گیاه گلدهی طبیعی نخواهد داشت.
لاله واژگون از گلهای بهاری است که به طور طبیعی در ماههای فروردین و اردیبهشت گل میدهد و گلهای آن بسیار جذاب و با عمر کوتاه است. اگر سوخها در دمایی غیر از آنچه که در محیط طبیعی دریافت میکنند، قرار گیرند ممکن است تاریخ گلدهی آن تغییر نماید.
لاله واژگون را از زیباترین گیاهان زینتی دنیا میدانند که بازارپسندی بالایی را دارا میباشد و بازارهای کشورهای اروپایی و آمریکایی توجه خاصی به این گیاه نشان میدهند. این گل به دلیل دارا بودن ساقههای بلند و قوی میتواند به عنوان گل شاخهبریدنی به اقصی نقاط دنیا صادر شود. به علاوه شکل و رنگ و اندازه گل در لاله واژگون بسیار زیبا و مناسب برای پارکها و باغها است و میتواند رقیبی جدی برای لالهها و سنبلهایی باشد که اکنون در تمامی نقاط جهان کشت و کار میشوند. لاله واژگون قابلیت فروش به صورت گیاه گلدانی را نیز دارا است.
لاله واژگون F. persica و F. imperialisاز مهمترین گیاهان صادراتی ترکیه به حساب میآید و بیش از ۱۰۰ سال است که به طور وسیع به بازارهای هلند و به میزان کمتری به بازارهای آلمان، انگلستان، آمریکا، ژاپن و دانمارک ارسال میشود.
امروزه هر عدد سوخ F. imperialis در فروشگاههای اینترنتی ۶ تا ۱۲ دلار به فروش میرسد.
اهمیت دارویی
لاله واژگون به دلیل داشتن خاصیت خلط آوری و ضدسرفه ، بیش از ۲۰۰۰ سال است که در طب سنتی چین مورد استفاده قرار میگیرد . ترکیبات شیمیایی موجود در لاله واژگون بهطور گسترده مورد بررسی قرار گرفتهاند که از جمله آنها میتوان به آلکالوئیدها، ساپونینها، ترپنوئیدها، استروئیدها، اسید سوکسنیک، تیمیدین و آدنوزین اشاره نمود. همچنین بنا به گزارشها، Fritillaria دارای فعالیت مهار تجمع پلاکت خون است. F. imperialis برای درمان بیماریهای مختلف از جمله گلو درد، سرفه، آسم، برونشیت، سل، غدد لنفاوی گردن، غدد تومور و سوزش ادرار استفاده میشود.
در بررسی ترکیبات شیمیایی سوخهای لاله واژگون، ۸ نوع آلکالوئید استروئیدی شامل آلکالوئیدهای Epeiedine،Ebeiedinone ، Isoverticine،Verticine ،Verticinone ،Hupehenine ، Ebeienine و Imperialineشناخته شدهاند که ترکیبات اصلی و فعال گونههای مختلف Fritillaria را تشکیل میدهند.
در مطالعهای تحت عنوان بررسی اثر ضد دردی عصاره آبی سوخ گیاه لاله واژگون در موش و مقایسه آن با مورفین مشخص گردید که عصاره آبی سوخ گیاه لاله واژگون دارای اثر ضد درد میباشد. تصور میشود وجود اثر ضد دردی این گیاه مربوط به حضور آلکالوئیدهای impericine و forticine باشد.
ازدیاد
Fritillaria به وسیله بذر نیز ازدیاد مییابد. در تابستان هنگامی که کپسول بذر شروع به قهوهای شدن کرد، قبل از باز شدن، بذور جمعآوری شده و در محل خشک و دارای تهویه نگهداری میشوند. بذرهای جمعآوری شده در پاییز کشت میگردند تا سوخ تولید شود. در این حالت در شرایط مناسب حدود ۵ تا ۶ سال زمان مورد نیاز است تا سوخی که توانایی تشکیل گل را دارا باشد ایجاد گردد. به علاوه گیاهان حاصل از بذر به دلیل دگرگرده افشانی گونههای Fritillaria شبیه والدین نخواهند بود.
مطالعات قبلی نشان داده است که F. imperialis را نمیتوان به طور سریع و موثر به وسیله روشهای سنتی مثل فلس برداری یا تقسیم سوخ ازدیاد کرد. استفاده از این روشها بهدلیل کم بودن تعداد فلس (۵-۳ عدد) به ازای هر سوخ و در نتیجه محدود بودن سلولهای مریستمی، دارای نرخ ازدیاد محدودی است.
پس از جدا نمودن فلس از قسمت های اصلی میتوان آن را درون خاک قرار داد. با این روش، هر جوانه موجود در فلس به سوخ تبدیل میشود. فلسها میتوانند مستقیماً ریشه بدهند، اما اگر به مدت ۶ هفته در ماسه مرطوب، پیت یا ورمیکولیت مرطوب در دمای ۱۸-۲۰ درجه سانتیگراد قرار بگیرند، نتایج بهتری به دست میآید (هارتمن و همکارن، ۲۰۰۱).درصورتی که سوخها به صورت افقی برش داده شوند و با چاقوی مخصوصی خراش داده شوند قابلیت باززایی سوخچه افزایش پیدا میکند. در گونه F. imperialis سوخهای تجاری پس از ۳ سال به دست میآیند.
دوره خواب Fritillariaخیلی کوتاه است و دوره رشد گیاه میتواند از اواسط مرداد ماه شروع شود. به همین دلیل ضروری است که سوخها در اواخر تابستان یا اوائل پاییز کشت شوند، در غیر این صورت سوخها در انبار شروع به ریشهدهی مینمایند. در اروپای غربی گلدهی در بهار آغاز شده و گیاهان در تیرماه پیر میشوند. لاله واژگون دارای یک خواب کوتاه مدت ظاهری پس از خشک شدن اندامهای هوایی است و در اوائل پاییز رشد آن شروع میشود و ریشهها و جوانهها شروع به رشد مینمایند )دی هرتوگ و لی نارد، ۱۹۹۳(. کیفیت و اندازه گل در F. persica به طور مستقیم به محیط و وزن سوخ بستگی دارد.
سوخهای لاله واژگون را به طور معمول نمیتوان پیشرس کرد اما تسریع گلدهی گیاه گلدانی امکانپذیر است. معمارمشرفی (۱۳۷۷) اثر سرما، جیبرلین و تاریخ کشت در بهبود کمی و کیفی و پیش رس کردن را در F. imperialis مورد بررسی قرار دادند. نتایج این تحقیق نشان داد سوخهایی که تیمار یک ماه سرما را دیدهاند پیشرس شدن گلها در آنها شدید بود. همچنین تیمار یک ماه سرما نه تنها در صفات زایشی بلکه در صفات رویشی مانند تعداد برگ و ارتفاع بوته اثر داشت، تیمار جیبرلین به غلظت ppm200 گلدهی لاله واژگون را افزایش داد و سقط جوانه گل را به مقدار زیادی کاهش داد. همچنین جیبرلین باعث افزایش طول و عرض گلبرگها و تعداد گل و وزن سوخ و سوخچهها گردید. تاریخ کاشت لاله واژگون نیز در پیشرس شدن گل موثر بوده است. برای بهبود وضعیت رویشی و بالا بردن کمیت و کیفیت گلدهی و ارائه گل خارج از فصل، استفاده از ترکیب تیمار سرمایی و جیبرلین در زمان مناسب پیشنهاد شده است.
کشت درون شیشهای شامل کشت انواع بافتها، سلولها و اندامهای گیاهی تحت شرایط سترون و در محیطهای غذایی کنترل شده است. قابلیت باززایی بافتها و اندامها و توانایی ازدیاد آنها مدتهاست که شناخته شده است. طی قرنها، باغداران با قلمهزنی گیاهانی که دارای صفات مطلوب بودهاند، از توانایی ارزشمند باززایی، برای ازدیاد گیاهان بهره گرفته اند.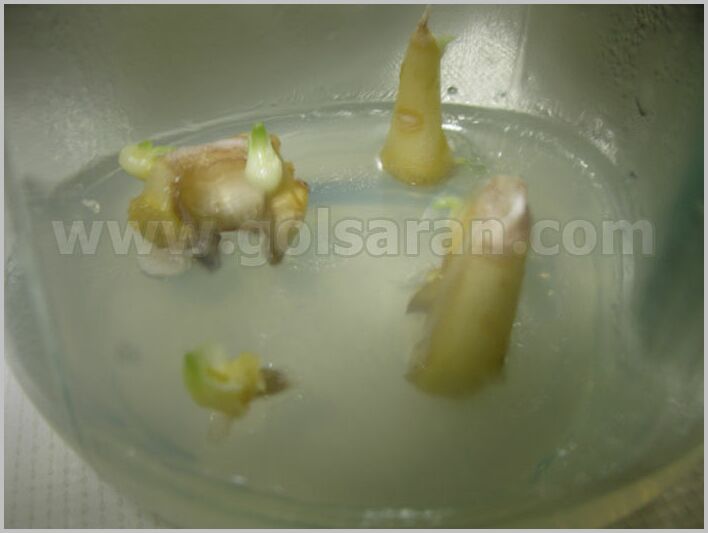
اولین آزمایش کشت بافت توسط هابرلند در بین سالهای ۱۸۹۸ تا ۱۹۰۲ انجام شد که حدود ۱۰ سال قبل از آغاز کشت سلولهای جانوری بوده است. هابرلند موفق شد سلولهای جدا شده از مزوفیل برگ را به مدت طولانی زنده نگه دارد ولی به دلیل این که محیط کشت او ساده و فاقد هورمونهای گیاهی بود، تقسیم سلولی در برگ صورت نگرفت. در اوایل قرن بیستم با پیشرفت روش گندزدایی و مشخص شدن نیاز به ویتامینهای گروه B و هورمون اکسین در کشتهای درون شیشهای، نخستین آزمایش موفق در زمینه القای رشد و تقسیم سلولی توسط وایت (۱۹۴۳) انجام شد. او ریشههای جدا شدهی گوجهفرنگی را در محیط گندزدایی شده مستقر کرد. تاکنون کشت درون شیشهای بیش از ۱۰ هزار گونه گیاهی با موفقیت انجام شده است.
از اولین تحقیق هابرلند که تلاش برای کشت بافت گیاهی بود، بیش از یک صد سال میگذرد. بعد از کشف هورمونهای گیاهی (اکسین در دهه ۱۹۳۰ و سایتوکنین در دهه ۱۹۶۰) و سنتز شیمیایی تنظیم کنندههای رشد گیاهی و متعاقب آن گسترش محیطهای غذایی در دهه ۱۹۶۰، آزمایشگاههای متعدد کشت بافت گیاهی در بسیاری از کشورها در دهه ۱۹۶۰ و ۱۹۷۰ تاسیس شدند. روشهای عمده ازدیاد گیاهان در شرایط کشت درونشیشهای شامل کشت جوانههای جانبی، تشکیل شاخه نابجا و جنینزایی سوماتیکی است. همچنین استفاده از این روش میتواند در زمینههای مختلف مثل نجات جنین، دورگ گیری بدنی، انتقال ژن، تولید گیاهان هاپلوئید، پلیپلوئیدی، ایجاد جهش و حفظ ذخایر ژنتیکی مفید باشد.
یکی از عوامل دیگری که در ازدیاد محصولات پیازی تاثیر میگذارد، کنترل خواب پیاز میباشد که هنوز به طور کامل مطالعه نشده است. در بسیاری از گونهها دوره خواب باعث میشود که گیاه قادر به تحمل شرایط محیطی شود. در عمل، تنظیم خواب در شرایط درون شیشهای مطلوب بوده و میتواند با اعمال تیمار سرمادهی پیازچه حاصل گردد.
گیاهانی که تحت شرایط درون شیشهای پرورش مییابند رشد سریعتری نسبت به حالت طبیعی دارند. در تحقیقی که روی کشت بافت گونه دارویی F. unibracteata صورت گرفته مشخص شده است که میزان رشد گیاهان حاصله ۳ تا ۵ مرتبه بیشتر از شرایط طبیعی میباشد.

کربوهیدراتها جز بسیار مهمی در هر نوع محیط کشت است. نیاز به کربوهیدراتها در شرایط درون شیشهای اولین بار توسط گوتریت در سال ۱۹۴۵ با کشت قطعات ریشه هویج مطالعه شد. از آنجا که معمولا در شرایط درون شیشهای، انجام فتوسنتز به علت نور پایین، تهویه ناکافی و رطوبت نسبی بالا بخوبی انجام نمیشود و یا اصولا به علت فقدان کلروفیل کافی در بافتهای کشت شده و یا قرار دادن آنها در تاریکی، فتوسنتز انجام نمیشود، لذا اضافه کردن منبع کربنی (هیدرات کربن) در محیط کشت ضروری است.
معمولا در کشت درون شیشهای از سوکروز یا قند معمولی به عنوان منبع تامین کننده کربن با غلظت ۱ تا ۵ درصد استفاده میشود، زیرا این قند توسط گیاه سنتز شده و به صورت طبیعی در گیاه منتقل میشود.
ریزنمونههای برخی از گونههای گیاهی روی محیطهایی که در آن از ساکارز اتوکلاو شده در مقایسه با محیط کشتهایی که در آن از ساکارز فیلتر شده استفاده شده بود رشد بهتری نمودند. این نشان دهنده این است که ساکارز در اثر اتوکلاو هیدرولیز میشود و در نتیجه سلولها قادرند از یک منبع گلوکز و فروکتوز آماده استفاده کنند.
نوع و غلظت قند انتخاب شده به نوع و سن مواد گیاهی در حال رشد بستگی دارد. در کشت پروتوپلاست سطح کمتری از کربوهیدراتها استفاده میشود ولی در کشت بساک به مقدار بیشتری از آن مورد نیاز است.
هیدراتهای کربن نه تنها به عنوان منبع کربن در متابولیسم سلول عمل میکند بلکه نقش مهمی در تنظیم فشار اسمزی محیط ایفا مینماید. با وجود نقش اسمزی منابع کربنی، در بیشتر پژوهشها تنها به جنبه غذایی آن پرداخته شده است در حالی که ترکیب اسمزی آن به میزان زیادی رشد اندام، بافت و سلول را تحت تاثیر قرار میدهد.
مواد معدنی مهمترین گروه مواد غذایی در کشتهای درون شیشهای پس از مواد قندی هستند. تعداد زیادی از نمکهای معدنی برای رشد و نمو گیاه لازم هستند. این نمکها به صورت پرمصرف و کممصرف طبقهبندی میشوند. بجز کربن، هیدروژن و اکسیژن، سایر عناصر ضروری که به مقدار نسبتا زیاد برای رشد و نمو بافتهای گیاهی مورد نیاز هستند، اصطلاحا عناصر غذایی پرمصرف (پر نیاز) مینامند. این عناصر شامل نیتروژن، فسفر، پتاسیم، کلسیم، منیزیم و گوگرد هستند که اغلب به صورت نمک به محیط کشت اضافه میشوند. میزان نیاز گیاه به این نمکها بیشتر از ۰/۵ میلی مول است. غلظت بهینه هر ماده غذایی برای دستیابی به میزان رشد بیشینه، به میزان زیادی بین گونههای گیاهی متفاوت است.
سلولهای گیاهی برای رشد طبیعی خود علاوه بر عناصر پرمصرف، در مقادیر بسیار جزئی به عناصر کممصرف نیز نیاز دارند. عناصر کممصرف در کلیه کشتهای گیاهان عالی شامل آهن، منگنز، روی، مس و مولیبدن هستند. میزان نیاز گیاه به این نمکها کمتر از ۰/۵ میلی مول است.
آهن مهمترین عنصر کممصرف است و به صورت کلات آهن (اتیل دی امین تترااستات آهن) یا FeEDTA مصرف میشود. کلات آهن با افزودن FeSO4 به Na2EDTA تشکیل میشود. غلظت آهن در محیط کشت ۱۰۰ میکرومول یا ۰/۱ میلیمول است. بنابراین مقدار مصرف آن با توجه به سایر عناصر کم مصرف، قابل توجه است.
بیشتر سلولهای گیاهی کشت شده قادرند همه ویتامینهای مورد نیاز خود را بسازند ولی این ویتامینهای ساخته شده کمتر از مقدار مورد نیاز برای این سلولهاست. بنابراین برای بهدست آوردن رشد بهتر بافتهای گیاهی اغلب لازم است که به محیطهای کشت یک یا چند ویتامین و اسیدآمینه افزوده شود. ویتامینها به عنوان کاتالیزور فرایندهای متابولیکی به کار میروند )نوری قنبلانی، ۱۳۷۱؛ خوشخوی، ۱۳۷۷(. ویتامینهایی که در محیط کشت بافت و سلول به کار میروند، شامل تیامین (B1)، نیکوتینیک اسید (B3)، پیریدوکسین (B6)، میواینوزیتول، فولیک اسید (M)، ریبوفلاوین (B2)، آسکوربیک اسید (C)، بیوتین (H)، توکوفرول (E) و پانتوتنیک اسید (B12) هستند. از میان این ویتامینها عموما تیامین ضروری تر است.
میواینوزیتول بهطور معمول در بسیاری از محلولهای ویتامیندار استفاده میشود. با آنکه میواینوزیتول یک کربوهیدرات است و ویتامین نیست ولی در مقدار کم باعث تحریک رشد سلول میشود. اعتقاد بر این است که میواینوزیتول به اسید آسکوربیک و پکتین شکسته میشود و در ساخته شدن فسفواینوزیتیدها و فسفاتیدیل اینوزیتول که نقش مهمی در تقسیم سلول دارد، شرکت میکند.
اکثر آزمایشهای کشت بافت روی بستری ثابت صورت میگیرد و بهطور معمول از یک عامل مولد ژل جهت این کار استفاده میشود. در کشتهایی که از محیط کشت مایع استفاده میشود، بافت غوطهور بیشتر به علت فقدان اکسیژن از بین میرود. برای جلوگیری از چنین حالتی محیط کشت را جامد یا نیمه جامد میکنند. آگار یکی از مشتقات نوعی علف هرز دریایی است که به صورت آماده وجود دارد و میتواند به عنوان ماده مولد ژل در اکثر محیطهای کشت استفاده شود.
آگار یک پلی ساکارید با وزن مولکولی بالا است که وقتی به محیط کشت بافت اضافه میشود، آب محیط را جذب کرده و یک حالت متخلخل و ژلهای به محیط میبخشد. در نتیجه تبادل گازها به ویژه اکسیژن و دیاکسیدکربن به راحتی صورت میگیرد. غلظت عادی آگار بین ۰/۶ تا ۰/۸ درصد است، اگر غلظت کمتر (۰/۴ درصد) به کار رود، محیط کشت خصوصا زمانی که pH محیط هم پایین است، خوب سفت نمیشود. اگر غلظت بالاتر (۱ درصد) انتخاب شود، در این صورت محیط کشت خیلی سفت میگردد و در نتیجه قرار دادن نمونه در محیط کشت را مشکل میسازد. افزودن ۰/۸ تا ۱ درصد زغال فعال، سفتی ژل آگار را کاهش میدهد. در بعضی موارد به جای آگار از موادی مانند فیتاژل، آلژینات، بیوژل و ژلرایت استفاده میشود. گاهی به جای آگار میتوان از پل کاغذی روی محیط کشت مایع نیز استفاده کرد.
میزان pHمحیط کشت تحت تاثیر ترکیبات (نمکهای غیر آلی، منبع کربوهیدراتها، عامل ژلی، زغال فعال و روش نگهداری) و اتوکلاو قرار دارد. میزانpH محیط بدون ریز نمونهها در طی نگهداری در اتاقک رشد به میزان ثابتی کاهش مییابد و این شاید به علت دهیدراته شدن محیط یا تهنشین شدن ترکیبات محیط همانند مواد معدنی باشد ولی علت اصلی آن به خوبی مشخص نیست.
عموما pH محیط کشت با NaOH یا HCl (1 تا ۱/۰ مولار) بین ۵ تا ۶/۵ قبل از اتوکلاو کردن تنظیم میشود. زیرا pHهای پایین (کمتر از ۴/۵) و pHهای بالاتر (بالاتر از ۷) عموما در کشت درون شیشهای باعث توقف رشد و نمو میشود. معمولا بعد از اتوکلاو کردنpH محیط کشت به اندازه ۰/۳ تا ۰/۵ واحد کاهش مییابد.
هورمونها نوعی ترکیبات آلی هستند که به صورت طبیعی در گیاهان عالی ساخته میشوند و روی رشد و نمو اثر میگذارند. هورمونها معمولا در نقاط مختلف گیاه فعال هستند. علاوه بر این ترکیبات طبیعی، ترکیبات شیمیایی مصنوعی نیز وجود دارند که همانند انواع طبیعی عمل مینمایند. مجموع هورمونها و ترکیبات مشابه شیمیایی تولید شده، تنظیم کنندههای رشد نامیده میشوند.
پنج گروه از تنظیم کنندههای رشد گیاهی در کشت درون شیشهای از اهمیت زیادی برخوردار هستند. این تنطیم کنندهها شامل اکسینها، سیتوکنینها، جیبرلینها، اسید آبسیزیک و اتیلن هستند. مهمترین تنظیم کنندههای رشد در کشتهای درون شیشهای، اکسینها و سایتوکنینها هستند به طوری که نسبت اکسینها و سایتوکنینها در محیط کشت، میتواند روند ریشه زایی و شاخهزایی را تحت تاثیر قرار دهد.
نور صرف نظر از فراهم کردن منبع انرژی برای فتوسنتز اثرات متعددی روی رشد گیاه دارد. مقدار کربوهیدراتها در شدتهای پایین نور یا تاریکی کاهش مییابد. تغییرات در مقدار هورمونهای داخلی یا سایر ترکیبات فیزیولوژیکی میتواند با تغییرات شدت، مدت و کیفیت (رنگ یا طول موج) نور تحت تاثیر قرار گیرد. این تاثیرات ممکن است در کشتها در هر مرحلهای روی دهد..
فعالیتهای فتوسنتزی در مراحل اولیه رشد گیاهان کشت شده در درون شیشه چندان مهم نیست ولی در مراحل بعدی مواد گیاهی کشت شده تا اندازهای برای اتوتروف شدن برانگیخته میشوند. نور برای فرایندهای مورفولوژیکی نظیر آغاز ریشهزایی و شاخهزایی و جنینزایی بدنی بسیار ضروری است. کیفیت نور، شدت نور و فتوپریود برای برخی کشتها بسیار تعیین کننده است. نور مناسب توصیه شده برای گیاهان ۳۵ میکرومول بر متر مربع در مجذور ثانیه به مدت ۱۲ تا ۱۶ ساعت در هر ۲۴ ساعت است. پیشنهاد میشود برای تامین این نور از لامپهای فلورسنت سفید استفاده شود. لامپهای آبی باعث تشکیل شاخه ها و نور قرمز القای ریشهزایی را در بسیاری از گونهها به همراه دارد.
دما از دیگر عوامل مهم در اتاقک رشد است. معمولا میزان درجه حرارت بین ۲۴ تا ۲۶ درجه سانتیگراد ثابت نگهداشته میشود و در بعضی موارد بسته به گونههای آزمایشی، دمای پایین (۱۸ درجه سانتیگراد برای گونههای سوخدار) یا دمای بالاتر (۲۸ تا ۲۹ درجه سانتیگراد برای گونههای گرمسیری) انتخاب میشود.
دمای مناسب برای رشد و نمو در محیط درون شیشهای معمولا ۳ تا ۴ درجه سانتیگراد بیشتر از دمای اتاق رشد است. در شرایط درون شیشهای میتوان با استفاده از دمای پایین باعث کند شدن رشد و نمو طبیعی شد و بدین وسیله امکان نگهداری مواد گیاهی در محیط درون شیشهای فراهم میشود.
رطوبت نسبی اتاقک رشد نیز از عوامل موثر در رشد ریزنمونه است. معمولا رطوبت نسبی اتاقک رشد بین ۴۰ تا ۵۰ درصد تنظیم میشود. از آنجا که رطوبت در داخل لولههای آزمایش نسبتا بالاست، رطوبت اتاقک رشد، احتمالا فقط روی تلفات آب از طریق لولههای آزمایش تاثیر میگذارد. رطوبت بالا در اتاقک رشد باعث افزایش درصد آلودگی خواهد شد و کاهش آن موجب از دست رفتن آب ریزنمونه میشود.
تهویه مناسب و تامین اکسیژن، عامل مهمی برای رشد سلولها و بافتها است (باقری و صفاری، ۱۳۸۳). به منظور جلوگیری از آلودگیهای کشتهای استریل، ممانعت از خشک شدن گیاه و محیط غذایی، کشت درون شیشهای در ظرف در بسته که تبادل گازی را بین اتمسفر ظرف و هوای بیرون را محدود میکند، انجام میشود. رشد گیاهان تحت تاثیر ترکیب محیط غذایی و اتمسفر گازی است. ترکیبات تولیدی توسط گیاه یا سایر بخشهای سیستم ممکن است در اتمسفر ظروف کشت تجمع یابد. مهمترین عامل موثر بر تجمع گازها، نوع و مقدار ماده گیاهی، خصوصیات فیزیکی ظرف و درپوش آن، محتوای محیط غذایی و وضعیت ماکروکلیما مانند دما، تهویه و شدت نور است.
اتمسفر محیط اساسا شامل ۷۸ درصد نیتروژن، ۲۱ درصد اکسیژن و ۰۳۵/۰ درصد دیاکسید کربن است. در طول فتوسنتز، گیاهان CO2 مصرف و اکسیژن تولید میکنند و طی تنفس، CO2 تولید و اکسیژن مصرف میشود. دیاکسید کربن و اتیلن و تعدادی دیگر از هیدروکربنها در طول کشت تجمع پیدا میکنند. با افزایش غلظت CO2، کاهش اکسیژن مشاهده میشود. البته گزارشهایی مبنی بر کاهش غلظت CO2 در اتمسفر محیط کشت بافت وجود دارد.
در سال ۱۹۷۴ سه مرحله ریزافزایی توسط موراشیگ شرح داده شد. مرحله اول شامل استقرار است که ریزنمونه در شرایط عاری از عوامل بیماریزا استقرار مییابد. مرحله دوم یا مرحله افزونگری مرحلهای است که شمار ریزنمونهها زیاد میشود. در مرحله سوم عادت دادن گیاهچه برای انتقال به خارج از لولههای آزمایش آغاز میگردد.
هدف اولیه از این مرحله دستیابی به درصد زیادی از ریزنمونههایی است که از عوامل بیماریزای سطحی عاری شدهاند. شرایط درون شیشهای مانند مقدار بالای مواد غذایی و ساکارز، رطوبت و دمای بالا که برای رشد گیاه مورد نظر مناسب است میتواند برای رشد میکرارگانیزمها نیز مطلوب باشد. به طورکلی گندزدایی سطحی در برگیرنده شستن بافت و در پی آن سترون کردن با یک یا چند ماده گندزدا میباشد. اساسا چهار منبع آلودگی محتمل است: گیاه (آلودگی داخلی و خارجی)، محیط کشت، هوا و بیدقتی در حین اجرای کشت. مهمترین منبع آلودگی ممکن است گیاه باشد، بنابراین مواد گیاهی بهلحاظ آنکه دارای طیف وسیعی از آلایندههای باکتریایی و قارچی است، باید به خوبی قبل از قرار گرفتن در محیط کشت کاملا گندزدایی شود.
برای کاهش آلودگی بهتر است ریزنمونه از اندامهای هوایی گرفته شود. از طرفی آلودگی قسمتهای داخلی گیاه کمتر از بخشهای خارجی آن است، بنابراین پوست سطوح خارجی حتیالامکان حذف و ریزنمونه از قسمت داخلی تهیه میشود. بهعلاوه هرچه ریزنمونه کوچکتر باشد احتمال آلودگی آن کمتر است.
برای حذف آلودگیهای سطحی ابتدا ریزنمونه باید با آب و صابون شسته شود تا گرد و غبار روی آن برطرف شده و سطح تماس ماده ضدعفونی کننده با ریز نمونه بیشتر گردد. پس از آن با شستن ریزنمونه در زیر آب جاری به مدت ۳۰ دقیقه تا ۲ ساعت به مقدار قابل توجهی میزان آلودگی ریزنمونههایی که از مزرعه گرفته شدهاند و همچنین بافتهای کرکدار، ریشهها و اندامهای ذخیرهای را کاهش میدهد. پس از شستشو، بافت ریزنمونه در یک محلول گندزدا فرو برده میشود تا آلودگیهای موجود در سطح آن از میان برود. برای ضدعفونی از الکل ۷۰ درصد برای چند ثانیه استفاده میشود. الکل ۹۶ درصد خیلی قوی است و باعث دهیدراته شدن شدید میشود. برای استریل کردن، نمونه به مدت ۱۰ تا ۳۰ دقیقه در هیپوکلریت سدیم یک درصد (NaClO) که دارای چند قطره توین ۲۰ (پلی سوربات) میباشد، قرار داده و سپس برای از بین بردن اثر هیپوکلریت معمولا سه بار به مدت ۲، ۵ و ۱۵ دقیقه با آب مقطر استریل شستشو میشود. توین ۲۰ برای حذف حبابهای هوا از مواد گیاهی و شکستن کشش سطحی بین آب و بافت گیاهی مفید است. دیگر گندزداهای مورد استفاده شامل هیپوکلریت کلسیم، پراکسید هیدروژن، نیترات نقره و کلرید جیوه است. برای افزایش تماس ریزنمونه با عامل ضدعفونی کننده بهتر است محلول حاوی ریزنمونه به طور مداوم تکان داده شود.
شرایط خاص درون شیشه باعث تشکیل گیاهانی با آناتومی، مورفولوژی و فیزیولوژی غیر معمول میشود. بعد از انتقال این گیاهان به شرایط خارج شیشه، این گیاهان ممکن است خیلی سریع به وسیله تغییرات ناگهانی شرایط محیطی نابود شوند.
قرار گرفتن تدریجی در معرض شرایط طبیعی منجر به سازگاری فیزیولوژیکی و مورفولوژیکی یعنی مقاوم شدن میگردد. از جمله این تغییرات، تغییر در خصوصیات اپیدرم، ضخامت برگ، تمایز مزوفیل برگ، تعداد و ساختار کلروپلاستهای برگ، کاهش یافتن تراکم روزنهها و تغییر شکل آنها از حالت گرد به بیضی است. همچنین کوتیکول برگها و میزان واکس آنها و میزان کلروفیل b در طول فرایند سازگاری افزایش مییابد. از نور کامل در طول مقاوم سازی باید پرهیز شود زیرا تنها قرار گرفتن گیاهچهها به مدت ۱ تا ۲ ساعت در برابر نور خورشید باعث مرگ آنها خواهد شد. از دیگر روشهای تسریع سازگاری استفاده از مواد ضد تنفسی مانند اسید آبسایزیک (ABA) و یا افزایش سرعت فتوسنتز با افزایش میزان CO2 است. اعمال سازگاری به ترتیب شامل مراحل زیر است: ۱) شستن ریشه و حذف تمام ذرات ژل (آگار)، ۲) انتقال ریزنمونه های ریشه دار شده به گلدان حاوی پیت: ورمیکولایت : ماسه به نسبت (۱:۱:۱)، ۳) آبیاری منظم با استفاده از یک افشانه دستی، ۴) نگهداری گیاهان در یک اطاقک مرطوب یا مکان دارای دستگاه رطوبتساز با شدت نور کاهش یافته (توسط پارچه یا تورهای مخصوص) در دمای ۲۵ درجه سانتیگراد، ۵) کاهش تدریجی رطوبت و افزایش شدت نور در طی چند هفته. در مواقعی هم لازم است گیاهان را توسط محلول های غذایی تغذیه نمود.
به طور کلی گزارشهای کمی در مورد کشت بافت لاله واژگون وجود دارد. عده ای محققان ابتدا تعداد ۲۲ عدد سوخ F. imperialis رقم روبرا ماکسیما را ۲ سال کشت نمود تا سوخچههای کوچک ایجاد نمایند. سپس جداکشتهایی را از قسمتهای مختلف آن تهیه کرد. درصد بالایی از جداکشتها آلوده و از بین رفتند و درصدی تولید کالوس نمودند. درصدی از کالوسها پس از ۲ ماه تولید سوخچه نمودند. همچنین با کاربرد جداکشتهای برگ، ساقه و نهنج اقدام به تکثیر این گیاه نمود اما درصد باززایی بسیار پایین بود.
در سال ۲۰۰۰ ویتومسکا تاثیر تیمار دمایی را بر ازدیاد درون شیشهای F. imperialisمورد بررسی قرار داد. او سوخها را ۸ هفته در دماهای ۴، ۲۲، ۳۰ و ۴۰ درجه سانتیگراد قرار داد، و گزارش کرد بهترین نتیجه از نگهداری سوخهای مادری در دمای ۳۰ درجه سانتیگراد بهدست میآید. همچنین تاثیر دمای اتاقک رشد نیز بر باززایی ریزنمونه ساقه مورد بررسی قرار گرفت. محیط کشت MS همرا با ترکیب هورمونهای رشد به میزان ۱ میلیگرم ۶ بنزیل آدنین (BA) همراه با ۰/۵ میلیگرم نفتالین استیک اسید (NAA) و دمای اتاقک رشد ۲۲ درجه سانتیگراد سوخچههای مناسبتری تولید نمود.
اوکاوا و همکاران نیز در سال ۱۹۹۸ کشت بافت F. camtschatcensis را در ۸ هفته و در دمای ۱۵، ۲۰ و ۲۵ درجه سانتیگراد مورد بررسی قرار دادند. بهترین شرایط برای تشکیل سوخچه در تیمار تاریکی همراه با ۰/۱ میلیگرم NAA در دمای ۲۰ درجه سانتیگراد گزارش شد.
الکساندرا و همکاراندر سال ۲۰۰۷ با مقایسه مقادیر ABA موجود در بافت مادری و توانایی باززایی آن در F. imperialis در شرایط آزمایشگاهی به این نتیجه رسیدند که در تمامی مراحل، بیشترین درصد باززایی از مواد گیاهی که دارای کمترین مقادیر ABA آزاد بود، بدست میآید.
پاککی و مورفی در سال ۲۰۰۲ درصد بالایی سوخچه از ریزنمونههای فلسی F. thunbergii به دست آوردند و این سوخچهها بعد از ۱۲ هفته از کشت، تولید برگ و ریشه نمودند. میانگین ۷/۱۷ سوخچه از فلسها درون محیط کشت MS حاوی ۱/۶۲ میکرومول NAA و ۴/۶۵ میکرومول کینتین بهدست آمد. کشتهایی که در چرخه ۱۶ ساعت روشنایی نور فلورسنت (۴۰ mol m–۲ s–۱) و ۸ ساعت تاریکی در دمای ۲۵ درجه سانتیگراد نگهداری شدند، نسبت به کشتهایی که در تاریکی مطلق در دمای ۲۵درجه سانتیگراد نگهداری شده بودند سوخهای مطلوبتری، تولید کردند.
در تحقیقی که روی گونه F. unibracteata به منظور تولید متابولیتهای ثانویه، صورت گرفت، زمانیکه جداکشتهای فلس در محیط دارای ۴/۴۴ میکرومول BA و ۵/۷۱ میکرومول IAA کشت شدند، مواد دارویی حاصل از سوخچههای بدست آمده بیشتر از شرایط طبیعی بود.
در پژوهشی محمدی ده چشمه در سال ۲۰۰۷ با عنوان جنینزایی غیرمستقیم از ریزنمونههای گلبرگ F. imperialis نتیجه گرفت که محیط کشت B5 حاویmg/l ۰/۱ BAP + mg/l ۰/۶NAA + mg/l ۰/۴IAA مناسبترین تیمار برای تولید سوخچه به تعداد ۵ عدد است.
شیا او و همکاران در سال ۲۰۰۰ تولید انبوه گونه F. hupehensis را همراه با سه نوع ریزنمونه در شرایط درون شیشهای مورد بررسی قرار دادند. بیشترین درصد باززایی از ریزنمونه فلس و پس از آن به ترتیب از ریزنمونه ساقه و برگ بهدست آمد. همچنین ۴ میلیگرم NAA باعث افزایش کالوسدهی ریزنمونه و بهبود ریشهزایی سوخچههای تشکیل شده گردید.
مشکل استقرار ریزنمونه لاله واژگون
بزرگترین مشکل استقرار ریزنمونه لاله واژگون در شرایط درون شیشهای حذف آلودگی است. آلودگیهای فوزاریومی، فیتوفترا و آلودگی باکتریایی از مهمترین موانع کشت بافت لاله واژگون هستند. از آنجا که فلسها در تماس با خاک هستند، گندزدایی آنها بسیار مشکل است. روشی که برای کنترل آلودگی در گیاهان سوخ دار همانند سوسن استفاده میشود، استفاده از تیمار آب گرم یا بخار آب گرم است. بر اساس پژوهش صورت گرفته در موسسه تحقیقات بیوتکنولوژی کشاورزی، این روش در مورد لاله واژگون کارایی نداشت و استفاده از جداکشتهای غیر از سوخ مورد تاکید قرار گرفت.
ویتومسکا و لوکازوسکا در سال ۱۹۹۸ چند روش گندزدایی را بر میزان آلودگی گیاهان و باززایی ریزنمونه های سوخ لاله واژگون مورد بررسی قرار دادند. آنها گندزدایی یک مرحلهای با غلظتهای مختلف کلرآمین T را با گندزدایی دو مرحلهای کلرآمین T و قارچ کش بنلیت مورد آزمایش قرار دادند. در این بررسی، گندزدایی با کلرآمین T درصد گیاهان سالم را به طور معنیداری افزایش نداد، در حالیکه استفاده از بنلیت درصد گیاهان گندزدایی شده را افزایش داد اما قابلیت باززایی، رشد و تشکیل ریشه در سوخهای استریل شده کاهش یافت.
در پژوهشی علت به دست نیامدن جداکشتهای سالم را آلوده بودن فلسهای گیاه لاله واژگون به آلودگی داخلی باکتریایی مربوط دانستند و عنوان نمودند که استفاده از سوخ این گیاه به عنوان جداکشت به دلیل وجود آلودگی داخلی حتی با اعمال تیمارهای مختلف از قبیل الکل، هیپوکلریت سدیم، کلرید جیوه و تیمار حرارتی میسر نمیباشد.
گونه های آسیایی و اروپایی لاله واژگون در مقایسه با گونههای آمریکایی دارای سوخهای بزرگ با تعداد بسیار کمتر فلس میباشند که بین ۴-۲ عدد متغیر میباشد.
گیاه لاله واژگون از جمله گیاهان در خطر انقراض کشور است که هرساله به دلیل برداشت غیرقابل کنترل گل و پیاز از رویشگاههای طبیعی، بیش از پیش در معرض نابودی قرار دارد. روش مرسوم ازدیاد از طریق کشت فلس به علت تعداد کم فلس و در نتیجه محدود بودن نقاط مریستمی و همچنین نیاز به مدت زمان طولانی به مدت ۵-۳ سال، دارای کارایی پایین است. ازدیاد بذری این گیاه نیز به ۷ سال زمان نیاز دارد. کشت درون شیشهای میتواند روشی مناسب برای ازدیاد انبوه گیاه لاله واژگون در زمان کوتاه باشد. در این روش علاوه بر ازدیاد سریع و جلوگیری از انقراض گونههای در معرض خطر، زمینه برای دستکاری ژنتیکی و اصلاح خصوصیات گل این گیاه و همچنین تولید متابولیتهای ثانویه از طریق کشت کالوس فراهم خواهد شد.
دانلود مقاله استخراج از پایان نامه اینجانب:
اثر تنظیم کنندههای رشد گیاهی در باززایی مستقیم سوخچه از ریزنمونه فلسی لاله واژگون
لطفا از مطلب لاله واژگون با درج نام سایت گلساران به عنوان منبع استفاده نمایید.